UMA NOVA UTILIDADE PARA O CIG
26/09/2020
Desde tempos remotos, o homem se empenha em exercer seu “domínio bíblico” sobre os animais, seja para prover sua alimentação, trabalho, transporte e até mesmo lazer. A passaricultura sempre foi uma paixão de muitas pessoas por todo o mundo e está essencialmente inserida na finalidade de lazer. Mais recentemente, também vem despertando um viés comercial, vislumbrando um negócio que talvez seja rentável.
Essa vertente econômica deflagra uma série de pesquisas, investimentos e estudos diversos. Na Europa já se deram ao trabalho de mapear o genoma completo de uma espécie, o diamante de gold. Por aqui, nos idos de 2000, passamos a ter a oferta deste serviço para algumas espécies de pássaros. O código de identidade genética – CIG -começou a ser ofertado com quatro (4) locos e atualmente atende padrões internacionais com mapeamento de quinze (15) locos de uma área do genoma que um dia foi chamada de “DNA lixo”, mas que exerce importantes funções na regulação dos genes.
Inicialmente, a utilidade principal era a aferição de paternidade, realizada por pesquisa mais simples entre o filhote e seu suposto genitor. Ocorre que, ao contrário do que acontece com os humanos, a passaricultura desenvolveu a técnica de promover cruzamentos endogâmicos visando fixar determinadas características tidas como desejáveis, e isso revelou que o sistema de aferição de paternidade por duo (filhote e genitor) não era suficientemente seguro, haja vista que parentes compartilham muitos alelos iguais e frequentemente laudos eram apresentados com equivocadas conclusões positivas. Assim, em 2014, passamos a contar com laudos de paternidade por trio (filhote, pai e mãe), muito mais seguro, mas que também pode apresentar falhas no caso de uma dose ainda maior de endogamia nos animais envolvidos.
A solução para esse novo obstáculo repousa na ampliação do número de locos pesquisados e, principalmente, na qualidade desses locos. Pesquisas são caríssimas, demandam o processamento de milhares de amostras, contratação de pessoal para essa pesquisa, organização de dados, cálculos de frequência, e, no final, pode ser que o locos aberto não tenha nenhuma contribuição a dar para essa finalidade de aferição de paternidade... É preciso que o locos abrigue uma maior diversidade de alelos, impondo uma baixa frequência na base de pesquisa, o que eleva o índice e torna mais seguro o cálculo. São algumas dezenas de milhares de reais, meses a fio, e nenhuma garantia da real utilidade de tal pesquisa para este fim.
Essa costuma ser uma dúvida frequente no meio passarinheiro: por que não abrem mais locos? Não é tão simples... Outra, igualmente frequente, é: por que os laboratórios não unificam os laudos?
Quanto a essa segunda dúvida, é bem mais complexa, eis que envolve direitos de propriedade intelectual na forma de detecção dos STRs que geram os CIGS.
Temos muito a aperfeiçoar, temos problemas, mas estamos em evolução e essa nunca é rápida. A agilidade da pesquisa está diretamente ligada ao tamanho do interesse econômico envolvido, porque é preciso muito investimento para cada passo a ser dado nessa longa caminhada pelo conhecimento.
Não sabemos do amanhã, mas hoje já é muito melhor que ontem. Não adquirimos mais “estórias”, dispomos de uma técnica segura para certificar o produto que estamos adquirindo. Podemos até saber a semelhança genética entre dois indivíduos. Foram duas décadas em acelerada evolução. Hoje já falamos com naturalidade e segurança em genética, mesmo que muitos ainda confundam a biologia com a matemática, ansiosos por respostas exatas. Não é área de exatas, é ciência biológica e nunca será exata!
O CIG também não é um fim em si. Não é um rascunho do diploma de campeão que muito sonham. É mais uma ferramenta para seleção, mais um parâmetro para criadores, mais uma informação para biólogos persistirem nos estudos.
Para os pássaros competidores, precisamos também estar atentos a todos os outros fatores, tais como: a) a descendência direta de exemplares de alto nível, lembrando que parentes colaterais na maioria das vezes herdam combinações diferentes, o que resulta em qualidades diferentes, por ser a fibra uma característica poligênica. O irmão do pássaro famoso não vai transmitir para os seus filhos as qualidades que o tio deles tem, se ele, irmão, também não as tiver herdado dos pais; b) a morfologia ou fenótipo do animal, eis que algumas variações são menos propensas a apresentarem as qualidades desejadas; c) o comportamento, que deve ser sempre ativo, esbanjando vitalidade; d) o histórico sanitário também é importantíssimo, pois uma doença, especialmente na infância, bem como algum transtorno psíquico experimentado, sempre deixam sequelas no animal, subtraindo-lhe uma parte do potencial genético; e) o manejo ofertado pelo criador é outro fator fundamental, aqui incluído o “casamento” do pássaro, e, por fim, não menos importante; f) a sorte de, na hora da marcação do seu pássaro, não sobrevir nenhuma circunstância desfavorável no ambiente do torneio.
E aqui, retornando ao tema das diversas utilidades que podemos extrair do CIG, gostaria de apresentar uma ideia que me ocorreu há alguns anos, a qual venho comentando desde então com alguns amigos e que penso poder contribuir para tornar menos aleatória a programação de cruzamentos. Trata-se da aferição de compatibilidade entre macho e fêmea, visando filhotes com uma “faixa” ou “banda” de similaridade já previamente visível em relação a um padrão predeterminado desejável.
En passant, tomo a liberdade de frisar algumas regras biológicas que já temos assentadas, onde o filhote herda metade do material genético do pai e a outra metade da mãe. Isso, em relação ao DNA nuclear. Também, importante destacar que, ao contrário do que pregam alguns precipitados, nenhum locos é mais importante que outro, até que saibamos a quais outros estão servindo em cooperação nessa poligenia, aos quais podem ou não estar ligados. Por fim, o traço logo após um número de alelo é apenas uma convenção e não quer dizer, necessariamente, que se trata de uma homozigose, embora realmente o seja na maioria dos casos.
Ao longo dos quinze (15) locos visíveis no CIG que nos é ofertado atualmente, os filhotes de um casal podem ter milhões de combinações possíveis em cruzamentos exogâmicos, mas podemos reduzi sobremaneira essa enorme quantidade de combinações na mesma proporção em que tivermos maior concentração de endogamia no plantel...
A ideia consiste em estabelecer um padrão desejado, escolhendo um pássaro-alvo que seja portador das qualidades que o criador deseja e que, preferencialmente, seja ancestral direto, em algum grau, dos exemplares que tivermos à disposição para reproduzir. Mirando no CIG deste pássaro-alvo, escolhemos o galador ou a matriz que desejamos usar para reproduzir. Então, dentre os do sexo oposto que tivermos à nossa disposição, podemos verificar a melhor e a pior similaridade da prole futura possível, estabelecendo o que chamei de “banda” ou “faixa” para decidirmos qual cruzamento privilegiar, qual macho é mais adequado para qual fêmea diante daquele objetivo predeterminado.

Conforme se nota e se lê, onde temos homozigoses aparentes as variações são reduzidas e isso “controla” melhor o padrão.
A melhor formação possível, nesse casal, resultará em filhotes com 95% de similaridade com o Peão; e a pior será similar em 63,3%. Essas margens de 63,3% e 95% são a “banda ou faixa”. A média da prole será 79,2% similar ao Peão.
➔ Casal 2. Usando o mesmo macho que tem similaridade de 75% com o pássaro-alvo e agora a fêmea 2, com similaridade de 81,7% com o pássaro-alvo. Entre eles, macho 1 e fêmea 2, a similaridade é de apenas 50%:
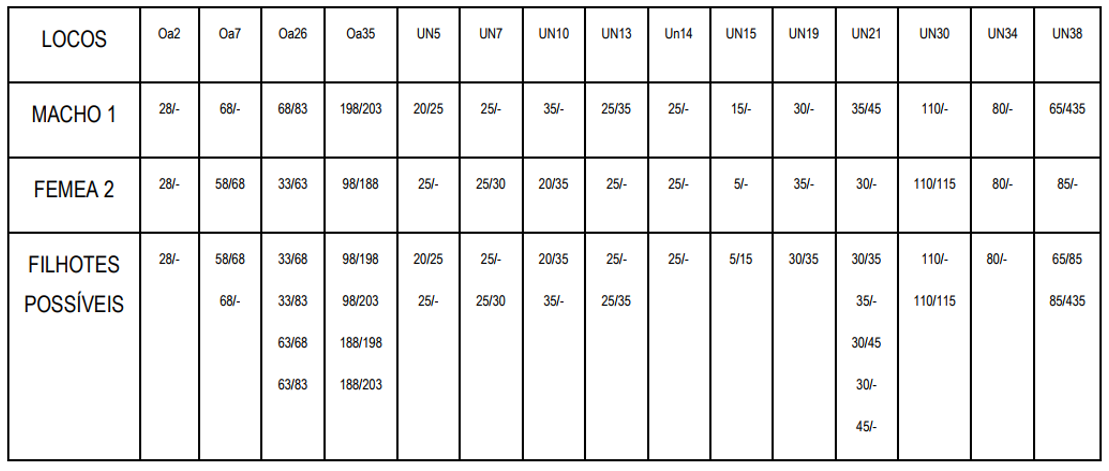
Observem que são cinco (cinco) as variações possíveis no locus UN21, demonstrando que biologia não segue um padrão puramente matemático. Outra questão a ser sublinhada é que embora a similaridade entre pai e mãe seja baixíssima, isso não afeta o conceito da compatibilidade, pois, conforme veremos abaixo, a média da prole deste segundo cruzamento foi ainda maior que a anterior.
Neste segundo cruzamento, a melhor formação possível resultará em filhotes com 98,3% de similaridade com o Peão; e a pior será similar em 65%. A média da prole será 81,65% similar ao Peão.
➔ Casal 3. Ainda utilizando o mesmo macho 1, que tem similaridade de 75% com o pássaro-alvo, para cruzar com a fêmea 3, que tem similaridade de 70% com o pássaro-alvo. Entre eles, macho 1 e fêmea 3, a similaridade é de 50%:
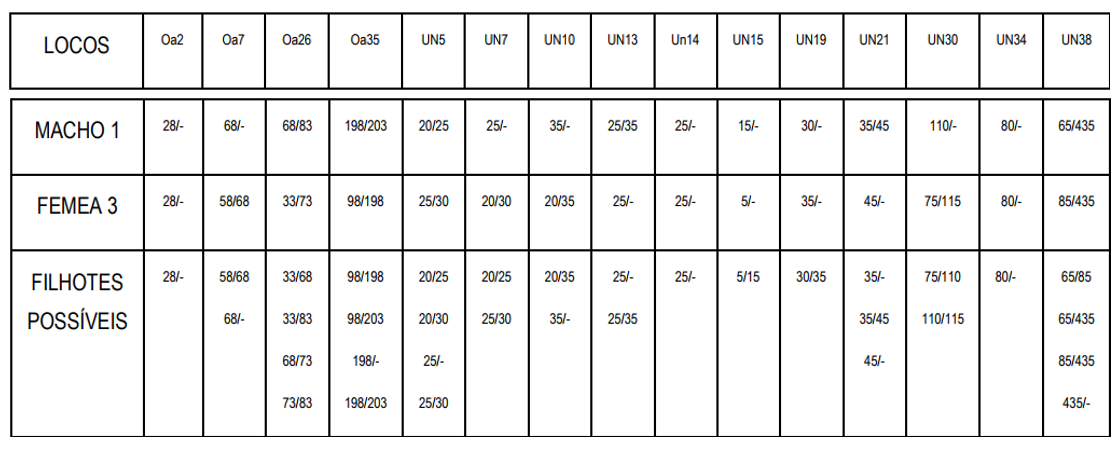
Neste terceiro cruzamento, a melhor formação possível resultará em filhotes com 93,3% de similaridade com o Peão; e a pior será similar em 56,7%. A média da prole será 75% similar ao Peão.
Cabe destacar algumas curiosidades explícitas nas três simulações acima:
a) embora as fêmeas 1 e 2 tenham a mesma similaridade com o Peão, reparem que a primeira é bem mais similar ao macho galador do que a segunda, e isso não influiu no resultado da prole, pois o exame de similaridade não se presta a programar cruzamentos, mas aferir o indivíduo que já existe, daí a relevância dessa análise de compatibilidade;
b) a terceira fêmea, que possui uma árvore mais fechada na linhagem, deu um resultado médio consideravelmente inferior às duas anteriores, apresentando uma faixa muito “larga”, indesejável. Seria mais indicado encontrar um outro macho, mais compatível, para cruzar essa fêmea, para não permitir que a prole seja tão heterogênea.
➔ Casal 4. Agora vamos usar um macho 2, que tem similaridade de 78,3% com o pássaro-alvo, para cruzar com a fêmea 3, que tem similaridade de 70% com o pássaro-alvo. Entre eles, macho 2 e fêmea 3, a similaridade é de 63,3%:
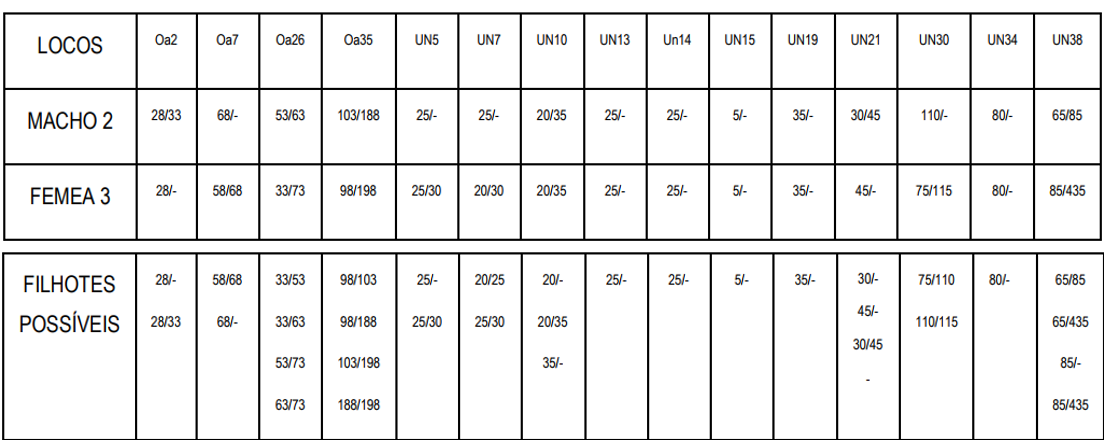
Neste quarto cruzamento, a melhor formação possível resultará em filhotes com 85% de similaridade com o Peão; e a pior será similar em 66,7%. A média da prole será 75,8% similar ao Peão. Nota-se que a faixa já foi reduzida à metade, menos de 20% de variação, e o piso da faixa subiu 10%. Outra curiosidade, é que a fêmea 2, utilizada acima, é filha desse cruzamento 4 e atingiu similaridade bem acima da média.
Certamente existem muitas outras utilidades interessantes para o CIG, essa aqui é apenas mais uma ideia...
Pontuo reafirmando que os pássaros têm diversos locos, cada um abrigando um par de alelos, que variam de pássaro para pássaro, daí o conceito de indivíduo; eles funcionam numa combinação que não sabemos, mas interligados certamente em razão do caractere poligênico da fibra; hoje nós só conseguimos visualizar quinze desses locos, abrigando trinta alelos e não podemos cometer o erro de resumir o resumo e ficarmos presos em análises superficiais em dois locos, só porque esses trazem maior variação. Cada locos é tão importante quanto o outro e, até que descubram a função de cada um, me parece que deveríamos dar mais atenção aos que não costumam variar. Mas isso já é assunto para outro texto...
Rodrigo C S Araújo
Gov. Valadares/MG
www.criadouroaraca.com.br - 2026 - Desenvolvimento:
![]()